
Transporte de gases respiratorios

Ingrese su ID de correo electrónico

Hb4 + 4 O2 <——-> Hb4 O8

Figura 2: Capacidad de oxígeno.
Cada gramo de Hb totalmente saturada puede unir 1,34 ml de O2 . Cada 100 ml de sangre contiene 15 gr de Hb. Luego cada 100 ml de sangre puede transportar 20,1 ml de O2 unido a la Hb más los 0,29 disueltos. En total: 20,4 ml de O2 /100 ml de sangre. Esta es la cantidad máxima de O2 que puede transportar los 100 ml de sangre para un hematocrito normal (O2 unido a la Hb y O2 disuelto para una PO2 determinada). Se denomina CAPACIDAD DE OXÍGENO. (Figura 2)
Por otra parte tenemos el CONTENIDO DE OXÍGENO que realmente está unido a la Hb.
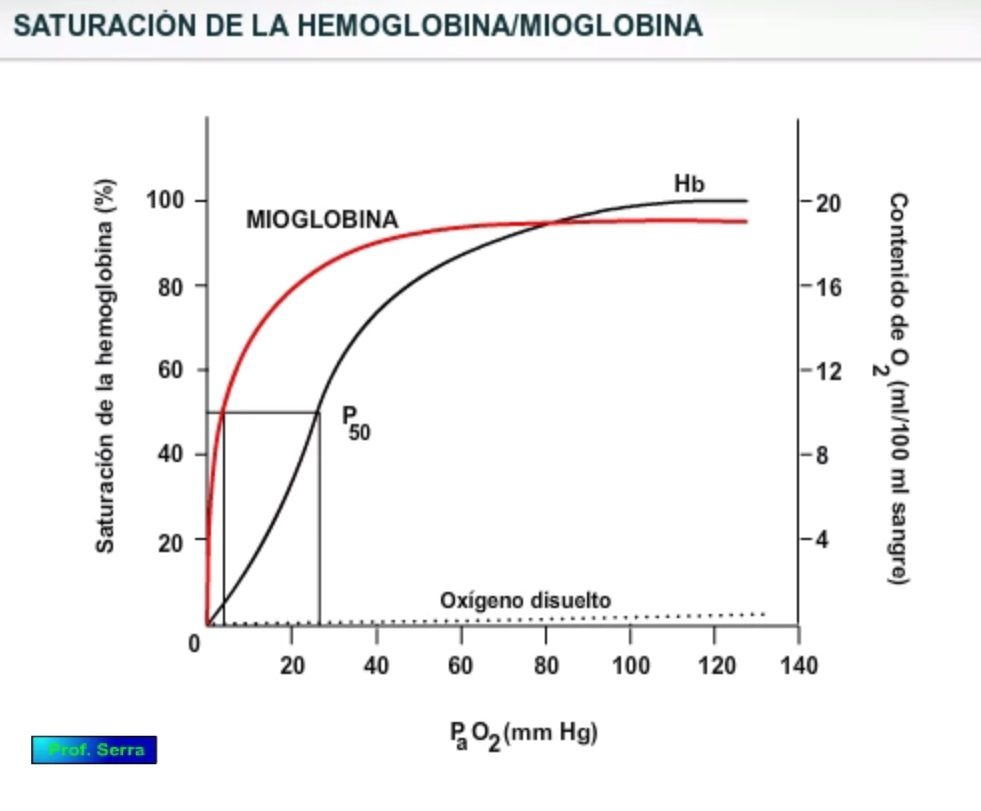
Si relacionamos ambas cantidades y la multiplicamos por 100, tenemos el PORCENTAJE DE SATURACIÓN DE O2 .

Figura 3: Saturación del oxígeno
% SO2 = (Contenido de O2 / Capacidad de O2 ) x 100








COEFICIENTE DE UTILIZACIÓN DEL OXÍGENO

Figura-13: Coeficiente de utilización del oxígeno.
Si medimos la cantidad de O2 en la sangre arterial (1,34 x [Hb](g/L) x %SO2) (19,8 ml O2 /100 ml sangre) y la cantidad en sangre venosa (14,8 ml O2 /100 ml sangre) y calculamos su diferencia, obtendremos la cantidad transferida en los tejidos que es de 5 ml de O2 /100 ml sangre. Si lo referimos al GC, será de 250 ml O2 /min, que es el consumo basal de O2 en los tejidos. Esta transferencia supone que en reposo sólo se transfiere el 25% de la capacidad total de saturación de la Hb y se denomina coeficiente de utilización.



EFECTOS DE LA VENTILACIÓN EN EL CO2 Y pH SANGUÍNEO
Modificaciones en la ventilación alveolar supone cambios tanto de la presión parcial de O2 como del CO2. Como se ve en la gráfica, si aumenta la PAO2 a valores altos, no se produce aumento de la saturación de la Hb una vez alcanzado el 100%, así mismo se necesitan caídas importantes de la PAO2 para que se ponga de manifiesto la hipoxia. Sin embargo el efecto es más directo sobre las variaciones del CO2 en sangre, pues éstas se reflejan rápidamente en cambios del pH plasmático.

Figura-17: Efectos de la ventilación en el CO2 y el pH sanguíneo.
Así tenemos que en el momento que se incrementa la ventilación alveolar y se produce una hipocapnia se observa rápidamente la aparición de una alcalosis respiratoria (lo que ocurre en las grandes alturas). La compensación viene dada por el riñón el cual actúa incrementando la excreción de bicarbonato. Por otro lado, cuando la ventilación alveolar es baja se produce un aumento del CO2 en sangre (hipercapnia) y en consecuencia se desarrolla una acidosis respiratoria, la cual si persiste se compensa por el riñón excretando ácido y preservando así el bicarbonato.
Por otro lado cuando se desarrolla una acidosis metabólica (hipoxia tisular, diabetes sacarina o tratada, ejercicio intenso, etc.) el sistema respiratorio compensa el exceso de ácido mediante un aumento de la ventilación. Y algo parecido ocurre pero en sentido contrario, cuando se desarrolla una alcalosis metabólica.
![]()
 Views Today : 25
Views Today : 25 Views This Month : 10194
Views This Month : 10194 Views This Year : 81015
Views This Year : 81015 Total views : 367078
Total views : 367078
Un comentario en “Transporte de gases respiratorios”