
Ingrese su ID de correo electrónico

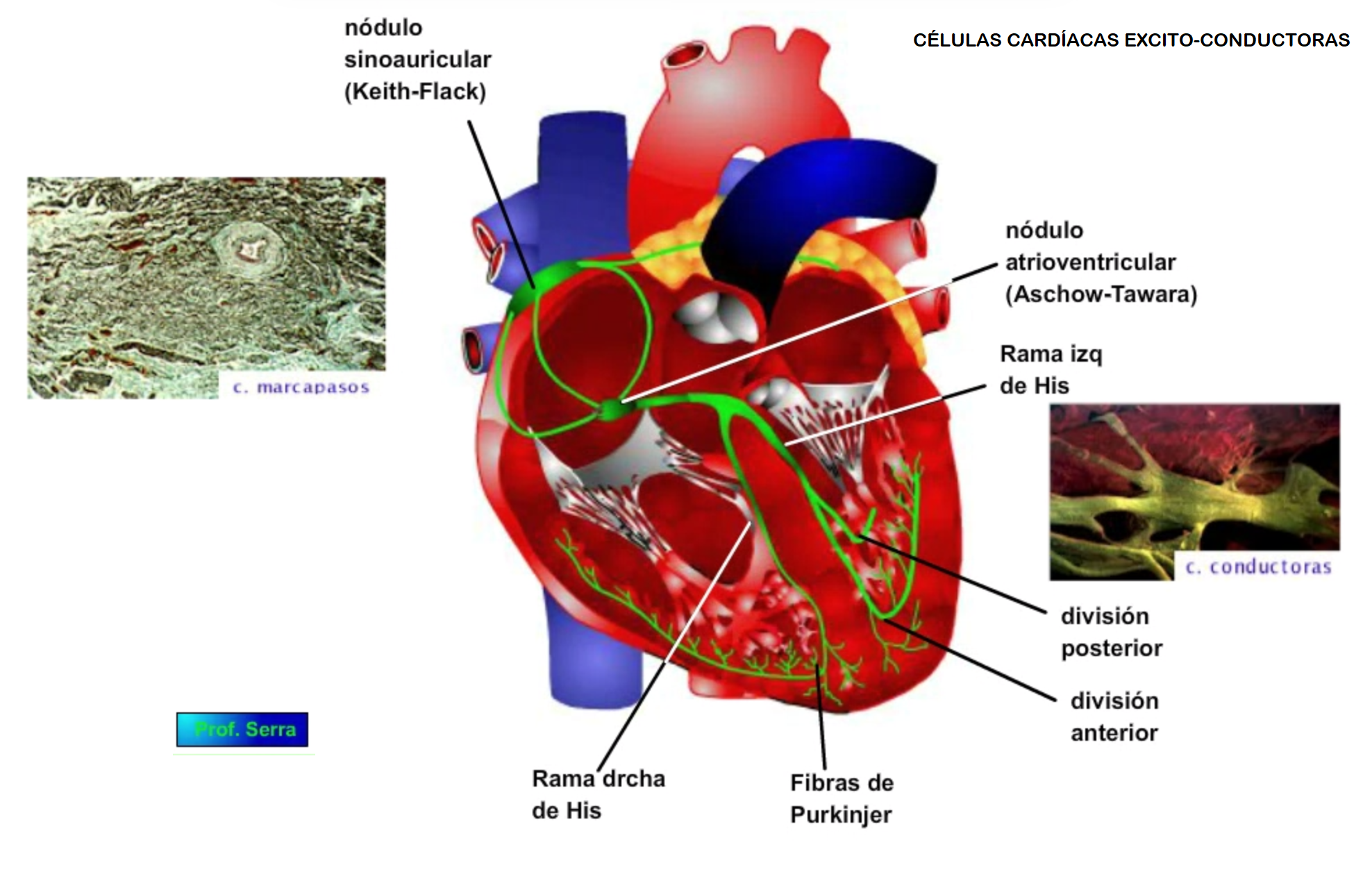


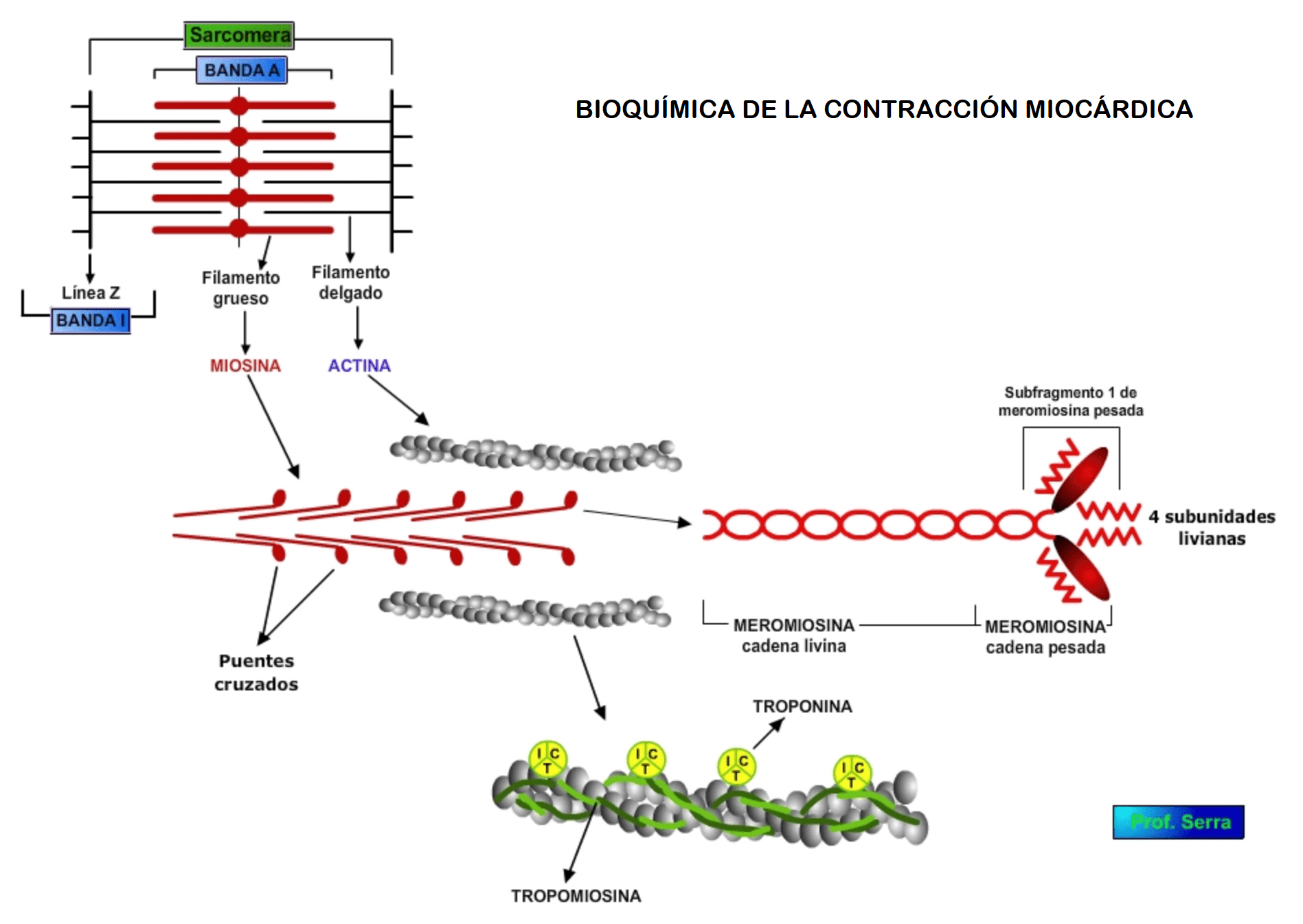



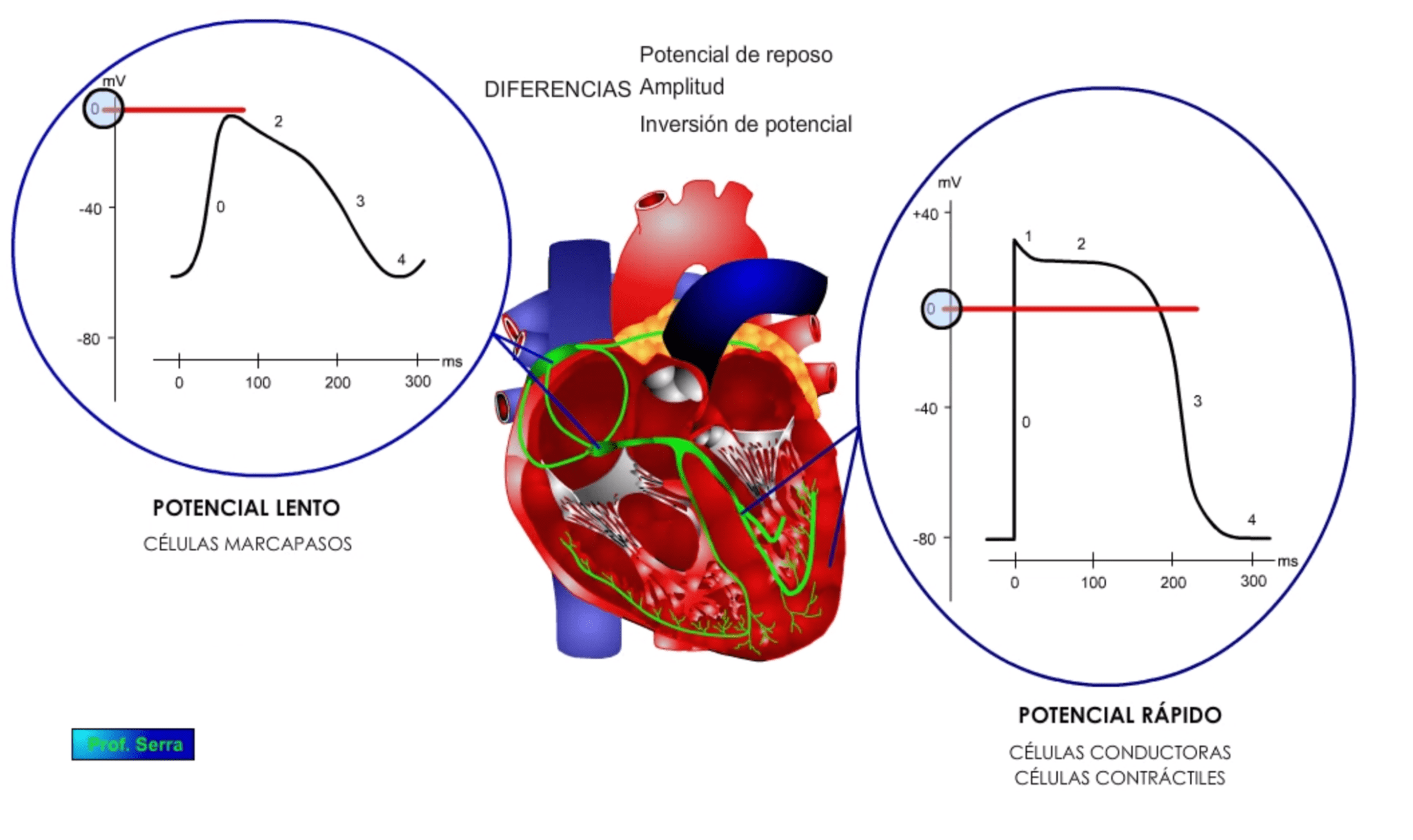






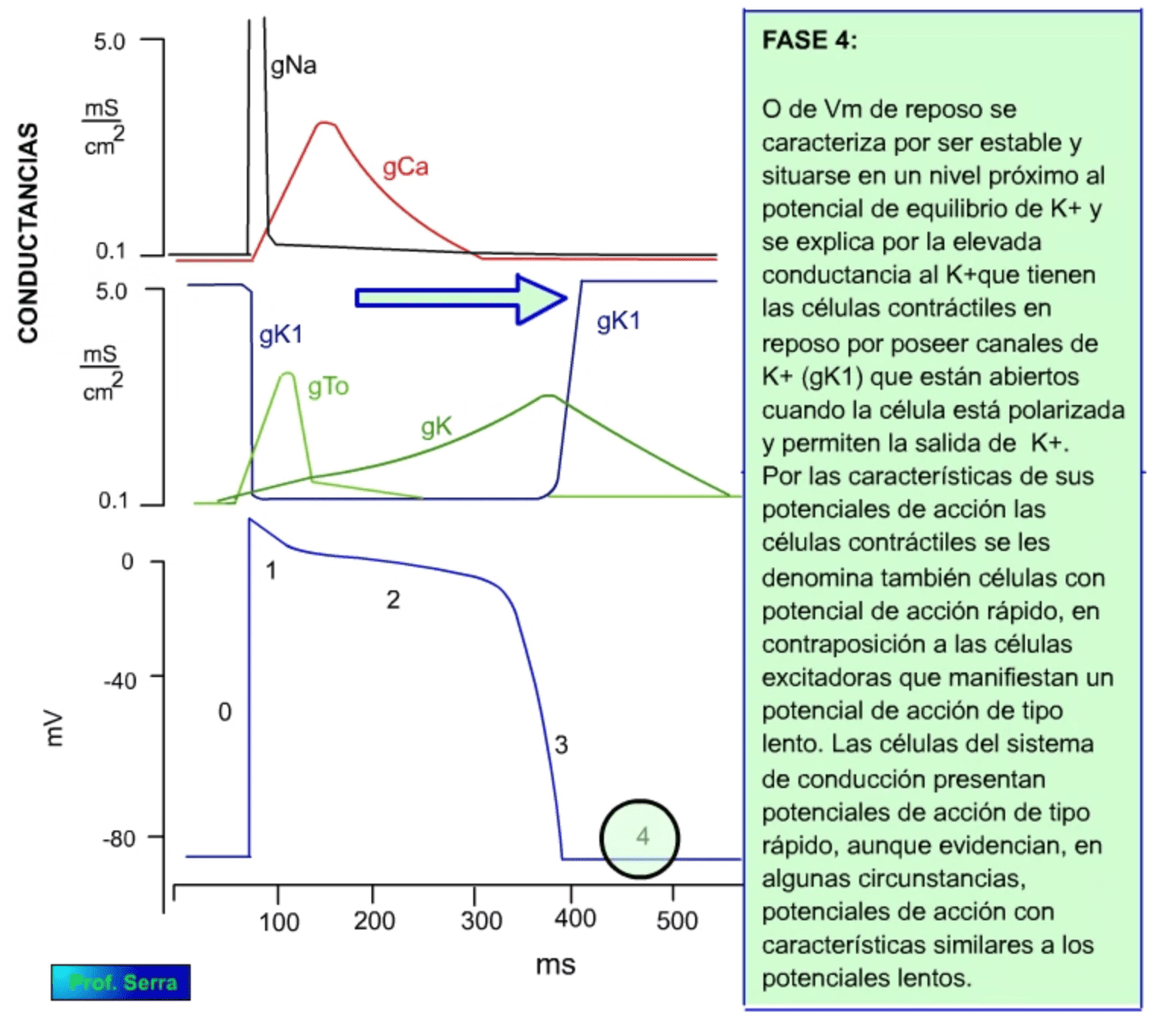
![]()
![]()
 Views Today : 580
Views Today : 580 Views This Month : 14930
Views This Month : 14930 Views This Year : 29708
Views This Year : 29708 Total views : 398855
Total views : 398855